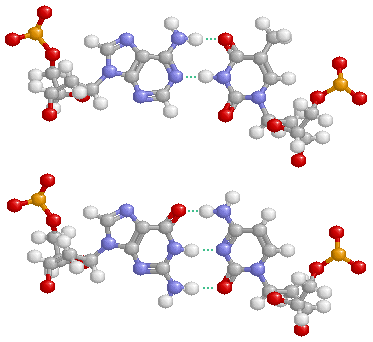
図1 すべての地球上生物の出発点であるDNA。遺伝情報を支える4種類の塩基(ATGC)と水素結合。
はじめに
2003年は,ワトソンとクリックがDNAの二重らせん構造を発表してから50周年にあたる上に,ヒトゲノムの完全解読もなされ,生命の不思議の鍵を握るDNAについて改めて考え直すきっかけの年になりました。
DNA鑑定,遺伝子組換え食品,遺伝子治療,DNAチップ,DNAコンピュータ,……。DNAに関する知見や技術は,単に自然科学的に大きな意味があるだけでなく,私たちのものの考え方や生活にも深くかかわってきており,将来的にもより幅広い領域で大きな意味を持ち続けるでしょう。
DNAは,図1のようにC,H,N,O,Pというたった5種類の元素からできており,有機化合物ですから熱やいろいろな薬品などによって容易に構造が破壊されたり分解したりしてその機能を失ってしまいます。つまり物質として見た場合には,極めて脆いとも言えるのですが,この地球上の多様で変化の激しい環境の中でそれを覆い尽くすほど広がってきたのは,その脆さをカバーするしたたかな生物システムがあったからなのです。
このページではその一端を,本サイトに掲載されている教材コンテンツで作成した画像を中心に紹介し,さらに生命の存在というものにも考察を加えてみたいと思います。
DNAを守るシステムとしてのセントラルドグマ
DNAは生物のシステムを機能させるための多数のタンパク質の設計図を有しており,さらにその設計図情報を遺伝情報として正確に(有性生物の場合は雌雄間でシャッフルして)子孫に伝えるという重要な役割を担っています。そのことは以下のようなセントラルドグマで示すことができます。
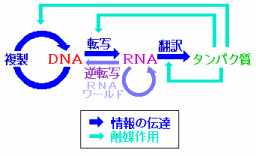
真核生物では,DNAは細胞内の核の中にあってタンパク質生産の現場に出て行くことはなく,mRNAに情報をコピーして生産現場に送り込むようになっており,このような手法もDNAの大事な構造を守るためのものと考えることもできます。
遺伝情報(DNAの塩基配列)とそれをもとに合成されるタンパク質の関係を示すのが遺伝暗号であり,これは高校の生物の教科書などにも載っています。
いろいろな攻撃に曝されるDNA
日本人の死亡原因の第1位は現在がんであり(国立がんセンターによるがんの統計参照),これはまさにDNAが環境中で様々な攻撃に曝されることによって引き起こされるものなのです。その要因としては以下のようなものがあげられています。
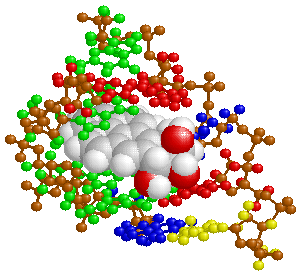
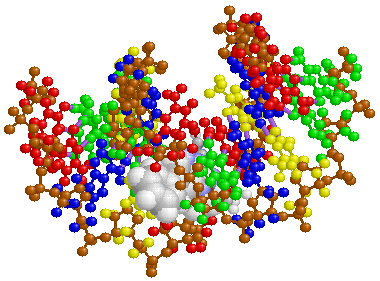
なお,ここで注意しておきたいのは,DNAは標準タイプであるB型自体がかなりの変形に耐えるほか,A型,Z型など細胞内でいろいろな働きをする時々に応じて多様な立体構造をとることも可能なことです。つまり,脆弱性の影に動的な柔軟性が潜んでいるわけで,これはシステムにとって極めて重要なことになります。
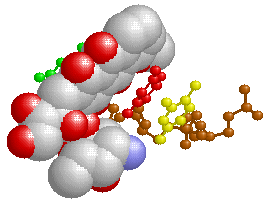
また,以上は正常細胞のDNAが攻撃された場合ですが,がん細胞のDNAを攻撃する抗がん剤の中には上と同様の機構を利用したものがあり,以下はその例になります。がん細胞だけ狙い撃ちできればいいのですが,正常細胞へも影響して副作用が出てしまいます。
DNAを守る仕組み
以上のように,DNAは様々な内的要因・外的要因によって損傷を受け,その設計図情報が撹乱されてしまいます。それでは生命システムを維持することができませんから,その影響を取り除いたり損傷したDNAを修復をするための仕組みなども多重的に備えているのです。
例えば活性酸素は,栄養物を体内で燃焼させてエネルギーを得るという過激な方法を採用している生物においては体内の化学反応では不可欠な物質であり,過剰な活性酸素が好ましくない影響を及ぼすのはもともと起こりうることなので,酵素や抗酸化ビタミン(C,E,β-カロチン等)などの抗酸化物質によってその影響を取り除くような機構が存在します。
下図は抗酸化能を持つ酵素の例です。
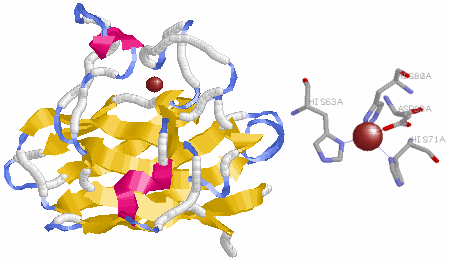
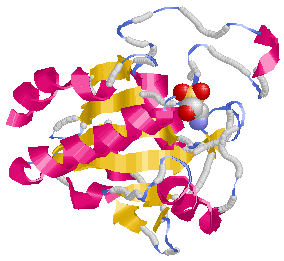
さらに,がん抑制遺伝子という頼もしい存在があることも忘れてはなりません。多くのがん細胞ではこのp53遺伝子が変異あるいは欠損しているので,正常なp53遺伝子を供給したり変異p53タンパク質を正常化したりすることができれば治療に結びつくと考えられています。
なお,酵素など高機能を有するタンパク質では,長い鎖中の離れた位置にあるアミノ酸残基が協調して活性部位を形成していること,その配列順序を決めているのがDNAであることを再確認しておく必要があります。生物が利用している分子の世界では,多様なネットワークが重要な役割を果たしていることがわかります。
DNAの“野心”
DNAを守るために
さいごに
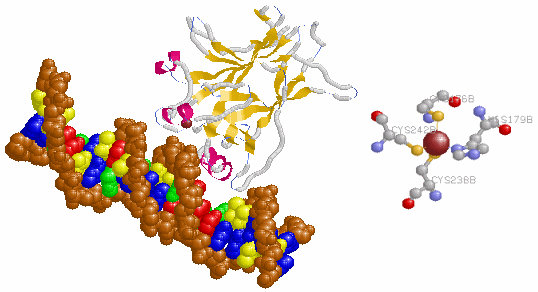
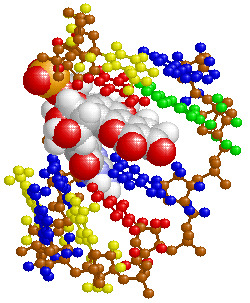
図10 がん抑制遺伝子p53の部分構造例(PDB1TUPのDNAとChain B)。
※右側は,zinc finger部分:Zn,Cys176,His179,Cys238,Cys242。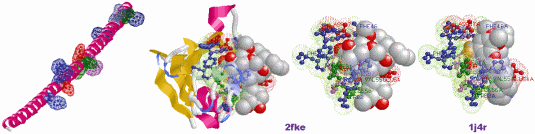
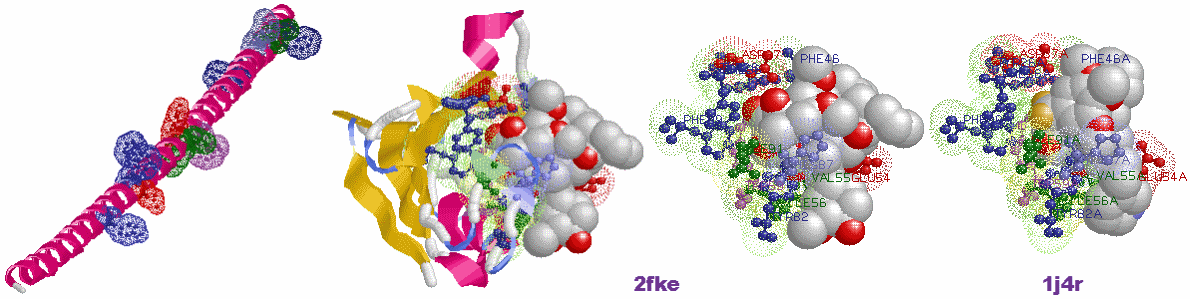
図11(拡大) タンパク質では,離れているアミノ酸が協調作業をして機能を示す(PDB2FKE・1J4Rを例に)。
左:2FKE・1J4Rと同じ下記アミノ酸配列ですべてα-helix構造にしたもの(MOLDA for Protein Modelingで作成)とSITE部位の強調表示。
GVQVETISPGDGRTFPKRGQTCVVHYTGMLEDGKKFDSSRDRNKPFKFMLGKQEVIRGWEEGVAQMSVGQRAKLTISPDYAYGATGHPGIIPPHATLVFDVELLKLE
中2つ:実際の2FKEとその活性部位(アミノ酸はすべてamino色表示,LigandはFK-506)。
右:2FKEの活性部位と同じ配列番号のアミノ酸をSITEとした1J4R(LigandはFKB-001)。
※参考:タンパク質の構築原理(理研ゲノム科学総合研究センター/タンパク質構造・機能研究グループ)
以上見てきたように,物質としては脆弱なDNAは同様に脆弱な有機化合物である様々なタンパク質や低分子を巧妙に利用して,地球という宇宙の中でも稀有なる揺籃のような環境の中でその存在を確たるものにしてきています(DNAにとっては細胞というものも“揺籃”に相当します)。その連係プレーはまるでスポーツ毎に決められている“人為的な”ルールのもとでのゲームの中でのできごとのようにも見えてきます。すべての生物がそのルールにしたがって共存と熾烈な争いを繰り広げ,突然変異や環境変化というルールの変更(フットボールからラグビーが生まれたように)やローカルルール(種差や個人差など)にも適応しながら,日々休むことなくそれぞれの営みを続けています。
脳の発達という戦略を選択した人間によって産み出された自然科学は,そのルールの解読はできてもなぜそんな競技を始めてしまったのかは説明できません。
ここで思い出すのは,この宇宙の中で物質が“存在”するためには,物質と光と観察者という三者が必要だという考え方です。つまり,物質世界が存在してもそれを認識できる知的生命体がいなければ存在していないことと同じであるという発想です。
同様なことが地球上の生命システムでも言えるような気がします。つまり,地球上の全生物はこれまでに見てきたようなルールにしたがっているけれども,そのようなルールが存在していることを知っているのは人間だけなのです(あるいは人間と違う手法でそのルールを理解している生物種もいるのかも知れませんけれど)。もしかしたらそれこそが宇宙の塵が集まってできた生命体のDNAがこの地球上に登場したときに持っていた“野心”なのかも知れません。
このような捉え方は突飛とも言えますが,そのように思索することで今人間が抱えている多くの問題を解決していく糸口が見つかるかもしれないと想定するのも,重要なことだと考えます。
このページで示されたことを踏まえていくつかおさらいしてみましょう。
上記 2,5 などから,安易に個々の生命システムを有する個体全体(個人)やその一部を故意に破壊してならないことは明白だと考えます。
次に 6 に関連するのが,近年研究が進められている遺伝子多型であり,これはある個人のゲノム解読が終了したことで遺伝子のルール解明の緒についたことを受けて,今後は個体差に注目していこうという流れの一つと見ることができます。その現況は例えば多型情報ネットワークやオーダーメイド医療実現化プロジェクト(文部科学省)で垣間見ることができ,今後のテーラーメード医療などへの利用が期待されています。ただし,遺伝情報は究極の個人情報であるため,その保護や活用についての開かれた議論によるルール作りが不可欠です。
また,同じく 6 に関連して想起したいのが,「ヒトゲノムと人権に関する世界宣言」の『A.人間の尊厳とヒトゲノム』に記載されている以下の2つの条文です。
第1条 ヒトゲノムは、人類すべての構成員が基本的に一体のものであること、並びにこれら構成員の固有の尊厳及び多様性を認識することの基礎である。象徴的な意味において、ヒトゲノムは、人類の遺産である。
なお,他の生物の多様な能力に依存する割合が最も高い人間は,この考え方をヒトのみならずすべての生物に対しても広めていくような基本的スタンス(実現の可否は保留するにしても)が要求されていると言ってもいいでしょう。
第2条 (a) 何人も、その遺伝的特徴の如何を問わず、その尊厳と人権を尊重される権利を有する。
(b) その尊厳ゆえに、個人をその遺伝的特徴に還元してはならず、また、その独自性及び多様性を尊重しなければならない。
以上のようなDNAに関する自然科学分野の知見(それは今後一層豊かになっていくでしょう)を踏まえた上での後段の記述は,これまでに読むことのできた生物と生命に関する多くの著作に拠っています。最近では,不断の真摯な思索によって『生命学』の確立を目指している森岡正博さんの実践的態度には教えられることが多く,あるいはこのページ作成自体もその感化なのかも知れません。ここでは,「生命学に何ができるか」p.426に記載されている『生命学の具体的作業』の項目を引用し,今後の本ページの加筆の指針にしたいと思います。
(1) 自己の問い直し、(2) 自分の人生における実験と検証、(3) 他者との出会い、(4) 生命世界の自分なりの解明と表現、(5) 得られた知見についてのコミュニケーション、(6) 社会変革への参画、(7) 先行者の表現物の学習、など
※今後取り上げたい注目キーワード:ロバスト,科学技術と社会(STS),……
【参考文献・Webページ】 その他の参考文献例はこちらをご覧ください。
Jmol版
→ でオリジナルPDBデータ表示(詳細はSummary Information参照)
PDB部分データによるコンテンツ集 | PDB部分データリスト | PDBデータのLigand結合部位
「分子の形と性質」学習帳 | 「生活環境化学の部屋」ホームページ
{kind=link}